Impact of elevated CO2 on aluminium tolerance of wheat in acid soils
Take home messages
- Response to elevated CO2 of Al-tolerant wheat genotypes appears to be affected by unidentified traits in the breeding parent lines and not by known carboxylate efflux mechanisms.
- The citrate and malate efflux wheat genotypes confer Al-tolerance in acidic soils and a greater overall tolerance is observed when citrate and malate efflux is combined.
- The ET8 wheat genotype showed superior performance under ambient CO2 rather than elevated CO2 for reasons that are not yet identified.
Background
Acidic soils (pH<5.5) are a major constraint to global agricultural production, often due to toxic levels of aluminium (Al) and manganese (Mn) and decreases in the availability of key plant nutrients, most commonly phosphorous (P) (Schroeder et al. 2013). Current estimates suggest that approximately 50% of the world’s arable land is acidic with 77-90 million hectares of topsoil in Australia alone currently acidic (Page et al. 2009). These estimates are predicted to increase in the future due to current farming practices leading to further soil acidification (Scott et al. 2000). While liming of acid soils is an effective ameliorant, it is not economically viable for farmers in developing countries or for ameliorating subsoil acidity (Kochian et al. 2004).
Of all the constraints under acidic soils, Al toxicity is considered the most important (Ma et al. 2001). When soil pH decreases below a pH of 5, Al3+ becomes soluble in the soil and its concentration exponentially increases as soil pH decreases (Kinraide 1991). Al3+ rapidly inhibits root cell growth and elongation and in the long term, decreases root cell division at the root apex (Ryan et al. 1993). This therefore results in poor water and nutrient uptake, which further exacerbates the problem of decreased nutrient availability in acidic soils. Aluminium tolerant plant species release carboxylates from the roots. These carboxylates such as malate and citrate can complex and thus detoxify Al3+ in the rhizosphere (soil directly in contact with the roots) (Kochian et al. 2004). Malate efflux in wheat is regulated by the TaALMT1 gene which is activated in response to Al3+, whereas citrate efflux is regulated by the TaMATE1B gene, which has a continuous efflux and should occur regardless of soil pH or Al3+ (Ryan et al. 2014).
In addition to these challenges, plants are being exposed to an ever changing climate especially in regards to CO2, water and temperature. The global atmospheric CO2 concentration pre-industrial revolution is estimated at ~280 μmol mol-1 and as of mid-2013 has increased to ~390 μmol mol-1 (Houghton and Ding 2001; Bala 2013). Climate models predict that global atmospheric CO2 will rise further to ~550 μmol mol-1 by the middle of the 21st century (Houghton and Ding 2001). The growth and productivity of plants are significantly affected by elevated CO2 (eCO2), however, the effects on root exudation and rhizosphere interactions is largely unknown (De Graaff et al. 2006; Jin et al. 2012).
This study aimed to examine the effect of eCO2 on Al tolerance of wheat genotypes that differ in malate (Al3+ activated) and/or citrate (continuous) efflux.
Methodology
A soil column experiment was conducted under ambient CO2 (aCO2) (~390ppm) and eCO2 (550ppm) in a FACE (Free Air CO2 Enrichment) system known as SoilFACE at the department of Environment and Primary Industries in Horsham, Victoria, Australia (36˚42’S, 142˚11’E). Wheat (Triticum aestivum L.) plants were grown in soil columns (60 cm long, 10cm in diameter), containing an acid Dermosol soil (pH~4.2). Six wheat genotypes differing in malate and/or citrate efflux were used: ES8 (-), Egret x Carazinho (Cz) (-), Egret x Cz (+Citrate), ET8 (+Malate), EGA-Burke x Cz (+Malate) and EGA-Burke x Cz (+Citrate & Malate). Of the six genotypes there are three isogenic pairs (differing in only a limited number of genes) which are ES8 (-) and ET8 (+Malate), Egret x Carazinho (Cz) (-) and Egret x Cz (+Citrate), EGA-Burke x Cz (+Malate) and EGA-Burkex Cz (+Citrate & Malate). Plants were harvested 108 days after sowing.
Preliminary results
Elevated CO2 significantly increased shoot biomass of EGA-Burke x Cz (+Malate) (24%) and EGA-Burke x Cz (+Citrate & Malate) (60%) (Figure 1). In comparison, eCO2 decreased shoot biomass of ET8 (+Malate) (-62%), also it did not affect shoot biomass of Egret x Cz (+Citrate) and Al3+ sensitive genotypes ES8 (-) and Egret x Cz (-) (Figure 1). There was no genotypic difference in shoot biomass between ambient and elevated CO2 treatments of Egret x Cz (+Citrate) and the elevated treatment of ET8 (+Malate) (Figure 1). All genotypes that released either malate and/or citrate had greater shoot biomass than corresponding genotypes that did not release malate and/or citrate (Figure 1). Greater shoot biomass was observed for EGA-Burke x Cz (+Citrate & Malate) compared to EGA-Burke x Cz (+Malate) for both CO2 treatments (Figure 1).
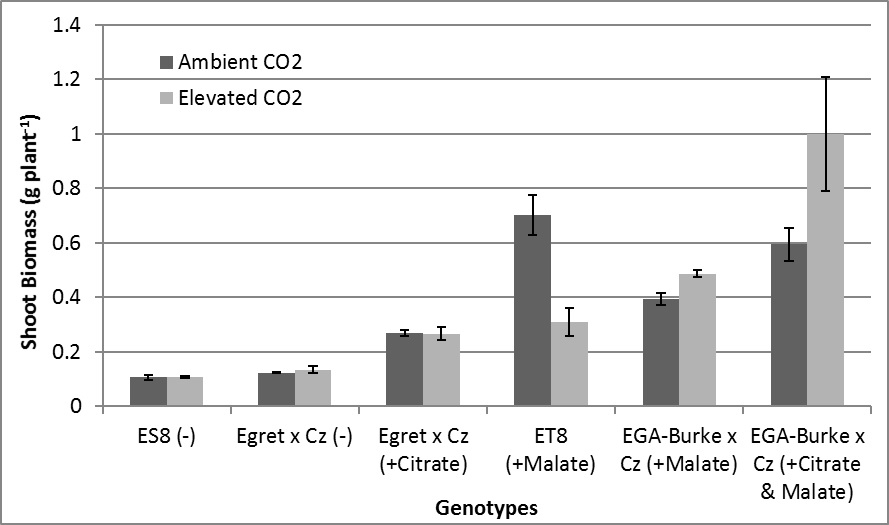
Figure 1. Shoot biomass of six wheat genotypes grown under ambient and elevated CO2. Cz = Carazinho, (-) = sensitive line, (+Citrate) = citrate efflux, and (+Malate) = malate efflux. Error bars represent ± standard error (n=4).
Similar to shoot biomass, root lengths observed similar trends with eCO2 significantly increasing the root length of EGA-Burke x Cz (+Citrate & Malate) (78%) (Figure 2). Additionally, decreases in root length were observed for ET8 (+Malate) (-62%) and no change for Al3+ sensitive genotypes ES8 (-) and Egret x Cz (-) (Figure 2). In comparison, eCO2 decreased root length of Egret x Cz (+Citrate) (32%) and did not affect root length of EGA-Burke x Cz (+Malate) (Figure 2). There was no genotypic difference in root length between the ambient CO2 treatment of Egret x Cz (+Citrate) and the elevated CO2 treatment of ET8 (+Malate) (Figure 2). All genotypes that released either malate and/or citrate had greater root lengths than corresponding genotypes that did not release malate and/or citrate (Figure 2). Greater root length was observed for EGA-Burke x Cz (+Citrate & Malate) compared to EGA-BurkeA x Cz (+Malate) for both CO2 treatments (Figure 2).
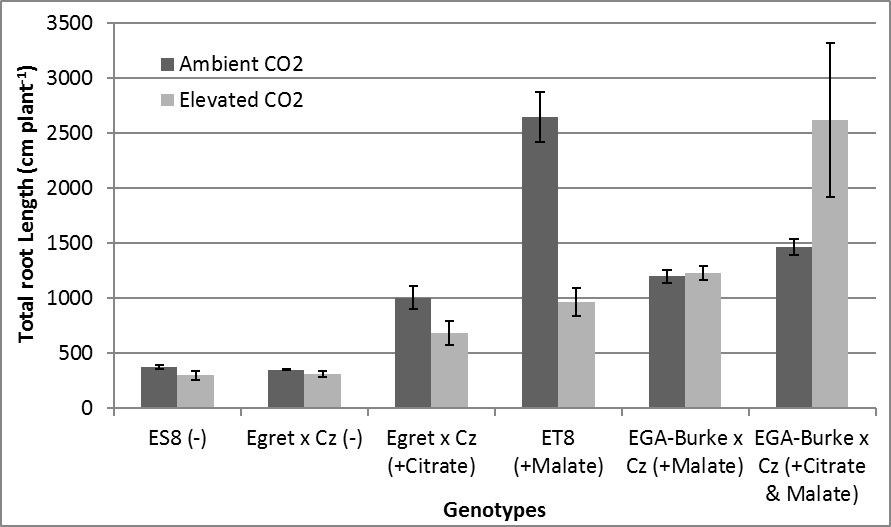
This initial study showed that the response of wheat to eCO2 varies significantly amongst different genotypes. Plants with carboxylate efflux (malate and/or citrate) displayed tolerance to acidic soils and in particular Al toxicity by recording significantly greater shoot biomass and root length compared to sensitive lines which had no carboxylate efflux. Following this study, further experiments will be conducted, in particular to try to determine why ET8 showed significant preference for aCO2 rather than eCO2 and to further confirm the greater combined tolerance of isogenic pairs citrate and malate compared to just malate. Ideally, this will lead to greater understanding and later proving beneficial to crop breeding and the selection of cultivars for acidic soil in an elevated CO2 environment.
Acknowledgments
This research is supported by Australian Postgraduate Award (APA) and Securing Food, Water and the Environment top-up scholarship. Thanks are also given to CSIRO Plant Industry for supplying wheat genotypes and the SoilFACE project for allowing access to the SoilFACE facilities.
References
Bala G (2013) Digesting 400 ppm for global mean CO2 concentration. Current Science 104: 1471-1472.
De Graaff MA, van Groenigen KJ, Six J, Hungate B, van Kessel C (2006) Interactions between plant growth and soil nutrient cycling under elevated CO2: a meta-analysis. Global Change Biology 12: 2077–2091.
Houghton JT, Ding Y (2001) IPCC-report. Cambridge University Press, Cambridge, UK.
Jin J, Tang C, Armstrong R, Sale P (2012) Phosphorus supply enhances the response of legumes to elevated CO2 (FACE) in a phosphorus-deficient vertisol. Plant and Soil 358: 91-104.
Kinraide TB (1991) Identity of the rhizotoxic aluminium species Plant-Soil Interactions at Low pH. Developments in Plant and Soil Sciences 45: 717-728.
Kochian LV, Hoekenga OA, Piñeros MA (2004) How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annual Review of Plant Biology 55: 459-493.
Ma JF, Ryan PR, Delhaize E (2001) Aluminium tolerance in plants and the complexing role of organic acids. Trends in Plant Science 6: 273-278.
Page K, Allen D, Dalal R, Slattery W (2009) Processes and magnitude of CO2, CH4, and N2O fluxes from liming of Australian acidic soils: a review. Soil Research 47: 747-762.
Ryan PR, Ditomaso JM, Kochian LV (1993) Aluminium toxicity in roots: an investigation of spatial sensitivity and the role of the root cap. Journal of Experimental Botany 44: 437-446.
Ryan PR, James RA, Weligama C, Delhaize E, Rattey A, Lewis DC, Bovill WD, McDonald G, Rathjen TM, Wang E, Fettell NA, Richardson AE (2014) Can citrate efflux from roots improve phosphorus uptake by plants? Testing the hypothesis with near-isogenic lines of wheat. Physiologia Plantarum 151: 230–242.
Schroeder JI, Delhaize E, Frommer WB, Guerinot ML, Harrison MJ, Herrera-Estrella L, Horie T, Kochian LV, Munns R, Nishizawa NK, Tsay Y, Sanders D (2013) Using membrane transporters to improve crops for sustainable food production. Nature 497: 60-66.
Scott B, Ridley A, Conyers M (2000) Management of soil acidity in long-term pastures of south-eastern Australia: a review. Animal Production Science 40: 1173-1198.
Contact details
Dominic Lauricella
Agricultural Sciences, La Trobe University, AgriBio – Centre for AgriBioSciences
5 Ring Rd, Bundoora, VIC
(03) 9032-7496
dlauricella@students.latrobe.edu.au
Was this page helpful?
YOUR FEEDBACK