Understanding nitrogen transformations in agricultural systems
Author: Lori Phillips, Cassandra Schefe, Roger Armstrong and Pauline Mele | Date: 28 Aug 2013
Lori Phillips, Cassandra Schefe, Roger Armstrong and Pauline Mele,
Department of Environment and Primary Industries, Victoria
Keywords: soil biology, mineralization, nitrification, denitrification, crop residues, fertilizer, crop rotation
Take home messages:
- Specific management practices alter the abundance and activity of microbes capable of processing both organic and mineral-N.
- There are direct links between the activity of microbes which process organic-N and those which process fertiliser; or mineral-N.
- These relationships can be manipulated to optimise N-use efficiency.
Background
Nitrogen (N) fertilizers are the largest variable input cost for grain production in Australia, and play a critical role in achieving high yields and premium quality grain. However, up to 50% of applied fertilizer N may be unavailable for crop uptake. Some N is lost from the soil by leaching and gassing off, while some is locked up in the soil organic matter in forms unavailable for plant use. Soil N-cycling, including the transformation of N from both fertilizer and soil organic matter into plant-available nitrate, is carried out by soil microbial communities (bacteria and fungi). Understanding which microbial communities are active in Australian soils during each step of the N cycle is the first steps towards improving N-use efficiency. Once we better understand the soil biological and chemical factors governing N-transformations in our agricultural systems, we can determine how agronomic practices can be used to synchronise N supply, from both fertiliser and soil sources, with crop demand.
Project overview
Soil N in agricultural systems is derived from two primary input types: fertilization and plant inputs via residue (shoots and roots of both legumes and non-legumes) decomposition. Once N enters the soil system, regardless of input type, it undergoes a series of microbial transformations which eventually results in the release of plant available NO3--N. These transformations can be generally divided into decomposition, depolymerization, mineralization, nitrification, and denitrification processes (Figure 1). Studies on the N cycle have typically focused on the nitrification process, but it is important to note that N-cycle transformations are not independent processes. The incorporation of plant residues into soil organic matter will both influence and be influenced by downstream N cycling processes. Products of one transformation become substrates in another, and any of the downstream products may be re-bound to soil organic matter via immobilisation processes. Thus, the dynamics of the system as a whole are inter-related. A primary goal of this project was to develop and use molecular tools to detect these N-cycling communities in Australian agricultural systems. Once we know who is there and what they are doing, we can then evaluate how to manipulate them to improve N-use efficiency.
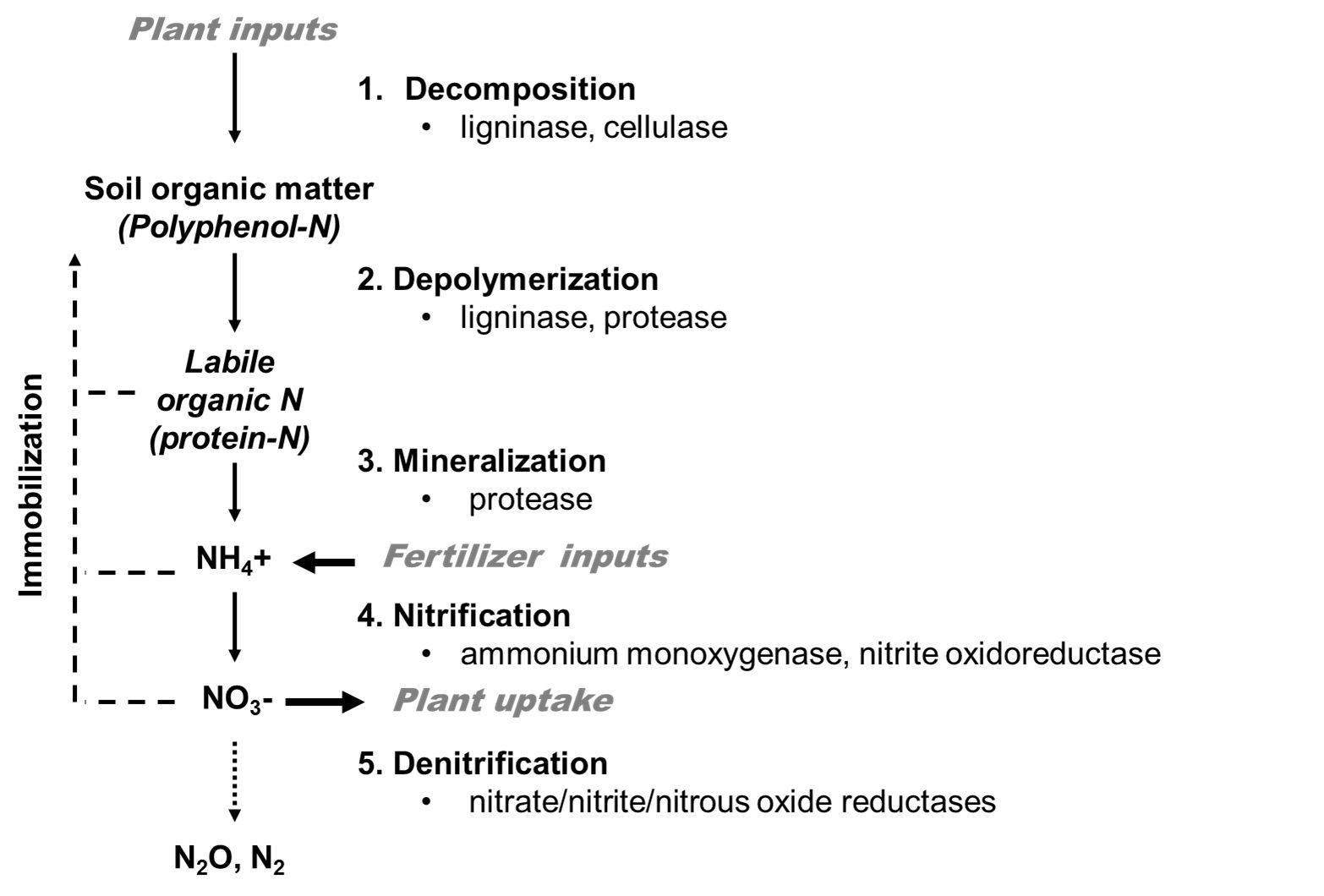
Figure 1. Microbially-mediating cycling of organic and inorganic nitrogen in agricultural soils.
Within DEPI-Victoria’s project, studies were designed to address the following specific questions:
- Can we apply molecular tools to identify key microbial communities active in the transformation of mineral and soil organic N to plant available N?
- Do previous crops influence microbial community functioning in subsequent cropping years?
- Do differences in the amount and quality of residue left behind impact microbial communities responsible for transforming organic and mineral N to plant available N?
- Can agronomic practices such as tillage be strategically used to manipulate N cycling microbial communities?
- Are specific N-cycling communities potential targets for new generation inhibitors?
Molecular tool development
Soil microbial communities use different enzymes (as listed in Figure 1) to process different compounds within their environment. Although a given community may possess the inherent genetic ability to, for example, nitrify ammonium to nitrate, the community may not be actively doing so at a specific point in time. To measure both the potential and the actual activity of microbial communities, this project developed DNA- and RNA-based measures of gene abundance and gene expression (genes code for enzymes). Genes for enzymes active in organic matter decomposition and depolymerisation (laccases, cellulases), mineralization of N from organic matter (proteases), nitrification (Archaeal and Bacterial ammonium monoxygenases, nitrite oxidoreductases), and denitrification (nitrite reductase, nitrate reductase, nitrous oxide reductase) were detected in all soils and protocols were developed to quantify their abundance (qPCR of DNA) and activity (RT-PCR of RNA). These protocols were then used to evaluate how agronomic practices, such as tillage, crop sequence, and residue management, impact the cycling of plant-available N.
Management impacts on microbial community structure and function
Two controlled environment studies and one intensive field sampling were used to assess how specific management strategies impact N-cycling microbial communities.
Rotation and tillage influence N-cycling
A controlled temperature and moisture study was used to determine which microbial communities were most closely linked to the release of plant-available N, and to evaluate how previous crops influence plant-available nitrogen at the start of the next cropping season. This study modelled the autumn break in soils from wheat, lucerne or green manure rotations. Soils from DEPI’s Sustainable Cropping Rotations in Mediterranean Environments (SCRIME) long-term trial site (Horsham, Victoria) were used in the study.
Summary results:
- The previous crop has a long-term impact on the structure and N-cycling activities of the microbial community present at the start of the next cropping season
- The abundance of N-cycling genes, including aminopeptidases, may be directly linked to soil N pools.
- Discing N-rich legume residues in, rather than leaving them on the surface:
- Primes microbial communities capable of breaking down or mineralizing organic nitrogen and subsequently cycling this organic N to plant available N
- Increases (more than doubles) the amount of nitrate-N which is released when soil moisture increases during the autumn break
Fertiliser and residue interactions influence N cycling
The importance of microbial communities that break down organic N sources was evident in a second controlled-environment study. This study was designed to investigate residue-fertiliser interactions on N-cycling communities in two different soils from the SCRIME (vertosol) and a DEPI-Hamilton (chromosol) field sites. Understanding the feedback mechanisms governing microbial use of both plant residue- and fertiliser-N also furthers our understanding how N is immobilized into organic matter.
Summary results:
- Different microbial nitrifying communities dominated in the two different soils, resulting in greater nitrification rates in vertosol soils than chromosol soils.
- Although the total N pools in the two soils were very different, the amount of mineral N (NO3 + NH4) present in each treatment in both soils was very similar, indicating microbial limits to the rate at which soil organic N pools are mineralized into plant available N
- Organic-N mineralization was inhibited by the addition of fertilizer N, which promotes the formation of microbial communities which predominantly nitrify fertilizer N rather than mineralize organic N.
- This inhibition is particularly evident at later sampling times, when both inorganic N and residue N should be maximally mineralized (Figure 2). In a field setting, these times would coincide with a time of high crop N need, as crops such as wheat enter the tillering stage.
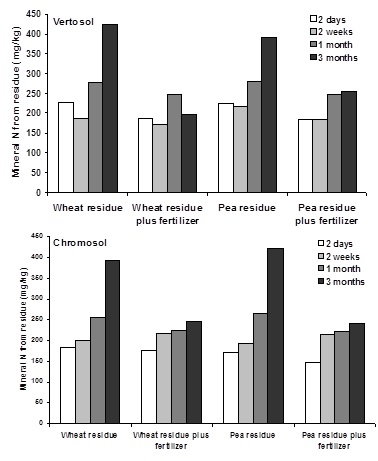
Figure 2. The addition of fertilizer decreases the release of plant-available nitrogen from soil organic matter and residues.
Field scale evaluation of management practices influencing microbial cycling of organic N
Long-term trials in the Wimmera region (DPI-Horsham SCRIME, vertosol) currently evaluate the impact of different agronomic management practices on plant health and grain yield. We used these existing trials to evaluate how management practices influence key N-cycling microbial communities throughout a cropping season. Soil samples were taken from 3 crops (wheat, pea and canola), under 2 tillage practices (zero and conventional tillage) and from 3 depths (0-5cm, 5-10cm and 10-20cm) at 5 key cropping stages (sowing, germination, tillering, anthesis and harvest).
Summary results:
Microbial community structure and function varied with crop, with tillage treatments, and with time. Although full analysis of the extensive generated data is ongoing, initial results are already providing insight into how specific management practices impact microbial cycling of soil organic nitrogen sources. Some of these results corroborate the findings of controlled environment studies, such as the finding that previous crops influence the nature of microbial communities present at the start of the next cropping season. Similarly, discing residues from a previous season into the soil further influences N-cycling microbial communities. For example, if the previous crop was a legume, then tillage stimulated greater numbers of microbes able to process organic nitrogen (Figure 3). As a result, tilled treatments coming out of a legume crop had 20-30% more mineral N available at sowing. The opposite was true following a canola or a wheat crop, where these organic-N cycling communities were greater in non-tilled treatments.
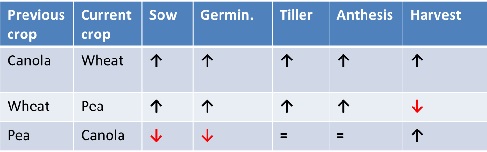
Figure 3. Relative abundance of microbes capable of processing soil organic nitrogen in non-tilled vs tilled systems. Up arrow indicates larger communities in non-tilled systems; down arrow indicates larger communities in tilled systems.
As found in the controlled environment studies, discing legume residues into the soil stimulates increased numbers of microbes able to process the N-rich residues. The obvious explanation for how this leads to increased mineral N becoming available is that more organic N-cycling microbes equals more mineral N. However, further investigation reveals that agricultural systems are not that simple. We need to dig a little deeper and understand what is actually happening to microbial communities when events such as tillage occur. This is where looking at the activity of microbes rather than simply their abundance provides additional insight.
In no-till systems at sowing, there is a significant relationship between the activities of microbes involved in N-cycling processes (Figure 4). These microbial activities are also significantly related to organic inputs and mineral-N release. These relationships are not found in tilled systems. One possible explanation for this is that tillage disrupts the soil biopore networks, which facilitate microbial movement and enhances microbial community linkages.
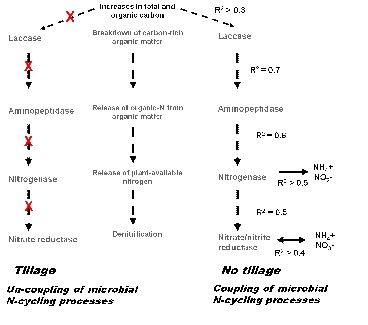
Figure 4: Relationship between the activities of different microbial N-cycling communities at sowing in tilled versus non-tilled systems.
If tillage disrupts the links between N-cycling communities, the question then arises as to why tillage treatments have higher mineral N production after a legume rotation? There are several possible reasons:
- While optimally you might want to keep this coupling intact, it is important to note that these interrelated processes also include denitrification processes.
- If mineral N is released from the residue too soon, then there is a greater likelihood of it being lost from the system, either via denitrification or through simple leaching of nitrate through the soil profile.
- These loss pathways may not be as significant following wheat or canola, as there is inherently less N in these residues.
- Although the microbial communities responsible for residue-N cycling are greater in abundance when legume residues are disced into the soils (ie Figure 3), they are not necessarily more active. Activity is actually lower in these tilled systems than in the no-till system (data not shown).
Project Summary and Management Implications
This GRDC-funded project has generated an unprecedented level of detail on how agronomy, soil chemistry, and soil biology interact to influence agricultural productivity. Our findings are extending our fundamental understanding of how agronomic practices influence soil microbiology, and how both may be managed to improve the synchronization of N supply with crop demand in Australian soils. Understanding which soil microbes are active in N-cycling at what time opens up new avenues for nutrient management.
These may include:
- Making informed decisions on paddock-specific tillage practices after specific crops
- The generation of new inhibitors which target alternate pathways within the nitrogen cycle
- Management of organic-N cycling communities is the key to optimizing organic N use and fertilizer efficiency
- Strategic timing of fertilizer application
- Specific to residue type from previous crops
Contact details
Lori A. Phillips
5 Ring Road, Bundoora, VIC, 3083
(03) 9032 7141
GRDC Project Code: DAV00106,
Was this page helpful?
YOUR FEEDBACK