USING ROTATION CROPS TO IMPROVE SOIL QUALITY
| Date: 08 Oct 2009
Soil quality issues are being researched within two crop rotation experiments that started in 1994 at Narrabri. They compare several crop rotations that include or exclude legume phases. The data presented here relate to the most recent 2-year cycles of these experiments. Following cotton harvest at the end of the previous cycle, rotation crops are sown (winter cereal, faba beans (grain) or vetch (green manured). Cotton is sown in the following October across all systems. The cropping systems also include continuous (annual) cotton in one experiment, with or without green-manured vetch each winter and a cereal-vetch treatment is included in each experiment.
Soil health has become more topical as some growers perceive a decline in their soil’s fertility and as soil-borne diseases impact on cotton cropping systems. Soil health relates not just to the presence or absence of the pathogens, but to soil characteristics that promote vigorous and healthy crop growth. Soil structure, the supply of nutrients and abundance of microbial communities can give some measure of the soils’ physical, chemical and biological properties.
Crop yield and N fertiliser response
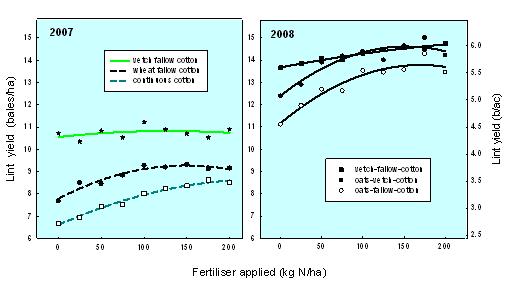
Initially, these experiments aimed to examine whether legume crops could supply enough N to enable farmers to reduce the amounts of N fertilisers needed to maximise lint yield. The responses to applied N can be seen in Figure 1. It quickly became apparent that the cotton yields we measured in the legume systems exceeded those gained from comparable non-legume systems. For example, less N fertiliser was required by the oats-vetch system because of the N input from the vetch, but importantly, the oat-vetch treatment had a significantly higher yield. The elevated lint yields in the oat-vetch system were consistent at all N fertiliser levels. This effect is due not just to the N input from the legume, but also to the improved soil environment created by growing the vetch. Our current aim is to evaluate these rotation crops and examine how they improve the soil environment.
Figure 1. Lint yield of the two rotation systems and the response to applied N fertiliser.
Soil Structure
Soil structural differences are measured with a cone penetrometer which determines the force needed to push a metal probe slowly into the soil to 600 mm depth. The results (Figure 2) indicate that the instrument penetrated the soil more easily in the legume systems compared with the non-legume systems. This effect was more evident in the subsoil. 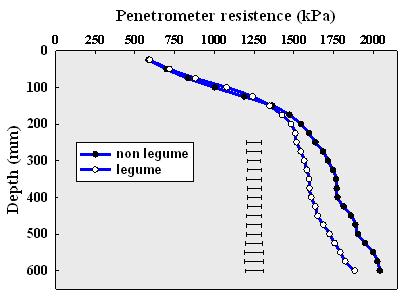
Figure 2. Resistance to inserting the cone penetrometer in to soil was lower following a legume (vetch) crop, compared with fallow.
This phenomenon has been observed in each year of the experiment. Soils that offer less resistance to root growth allows the crop to explore a larger volume of soil, thereby enabling the crop to take up more nutrients and to access more soil water.
Bulk density indicates how the soil components are packed together. The figure below (Figure 3) indicates the rotation crops reduced soil density, particularly in the subsoil. This can promote root growth and uptake of nutrients and water. The continuous cotton systems have much denser soil that those systems supporting rotation crops.
Porosity indicates how well aerated to the soil is, higher porosity allows greater root growth and can reduce N loss from denitrification. These soils tend to be less prone to waterlogging but may allow greater through drainage of water, nutrients and salts. There is more pore space in the soils that have had rotation crops, which promotes better soil aeration and water-holding capacity. 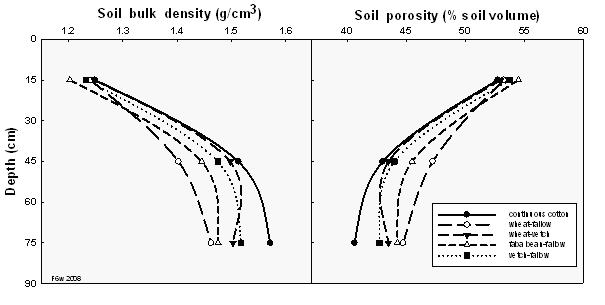
Figure 3. Soil bulk density and porosity (the volume of soil occupied by air and water) as affected by various rotation crops.
Nutrient availability
Cotton leaf nutrient concentrations at flowering
Nutrient content of the youngest mature leaf (5th uppermost leaf from terminal) was determined at first flower (Table 1). Leaves collected from cotton plants growing in the cereal-vetch systems contained slightly higher or similar concentrations of most nutrients than those in the cereal-fallow systems. Iron, manganese and sodium concentrations were slightly lower in the cereal-vetch systems where better soil aeration and good physical soil structure maintained these nutrients at levels conducive to healthy plant growth. High levels of these nutrients can occur in poorly-aerated soils.
Table 1. Leaf nutrient concentrations were normally higher where legumes were included in the crop rotation sequence.
|
Nutrient
|
cereal-fallow
|
cereal-vetch
|
|
Nitrogen (%)
|
4.60
|
4.62
|
|
Phosphorus (%)
|
2.99
|
3.16
|
|
Potassium (%)
|
1.60
|
1.66
|
|
Sulfur (%)
|
0.88
|
1.00
|
|
Calcium (%)
|
4.4
|
4.5
|
|
Magnesium (%)
|
0.73
|
0.77
|
|
Sodium (mg/kg)
|
839
|
798
|
|
Zinc (mg/kg)
|
23
|
25
|
|
Iron (mg/kg)
|
370
|
252
|
|
Manganese (mg/kg)
|
101
|
97
|
|
Boron (mg/kg)
|
49
|
49
|
|
Copper (mg/kg)
|
5.2
|
5.9
|
Nutrient uptake by the crop
At the end of flowering (cut-out), crop dry matter (DM) was determined by cutting 1 m sections of crop row and this material was dried, weighed, milled and analysed for nutrient content (Table 2). Cotton produced 10% more DM in the cereal-vetch systems than the cereal-fallow systems and took up greater quantities of most nutrients, including 33% more N, 7% more P, 12% more K and 23% more Zn. Sodium uptake was slightly higher in the oat-vetch system, but the amounts were low, compared with level of sodium taken up in sodic soils: cotton growing in more highly sodic soils may take up ten times the amounts reported here.
Table 2. Nutrient uptake by cotton in two crop rotation systems.
|
Nutrient
|
cereal
|
cereal-vetch
|
|
Crop DM (t/ha)
|
10.5
|
11.6
|
|
Nitrogen (kg/ha)
|
180
|
239
|
|
Phosphorus (kg/ha)
|
32.9
|
35.3
|
|
Potassium (kg/ha)
|
217
|
244
|
|
Sulfur (kg/ha)
|
54
|
57
|
|
Calcium (kg/ha)
|
211
|
228
|
|
Magnesium (kg/ha)
|
43
|
51
|
|
Sodium (kg/ha)
|
6.8
|
8.6
|
|
Zinc (g/ha)
|
145
|
178
|
|
Iron (g/ha)
|
1600
|
1400
|
|
Manganese (g/ha)
|
458
|
523
|
|
Boron (g/ha)
|
399
|
428
|
|
Copper (g/ha)
|
71
|
59
|
Soil micro-biology
Land management practices (such as crop rotation and tillage) can influence the abundance and composition of soil microbial communities. Microbial biomass is defined as the living component of soil organic matter. It has a much faster rate of turnover than total organic matter (OM) and thus can provide a more sensitive measure of changes due to land management practices.
Most nutrient transformations are mediated by soil microorganisms. The microbial biomass is responsible for decomposing crop stubble which is incorporated into soil OM. From there, it is further mineralized and nutrients released for crop nutrition. The soil microbial biomass itself can become a substantial store of nutrients which are recycled as these microorganism populations grow and decline. The rate at which soil organic N is mineralized can provide a useful integrator of chemical, physical and biological aspects of microbial community; it reflects both the decomposer activity of general microbial population and the activity of specific communities such as nitrifying microorganisms. Microbial activity (soil respiration), microbial biomass carbon (C) and N, and N mineralization potential are some of the measures suggested as potential indicators of biological status of soil.
We measured these microbial properties using surface soil samples (0-10 cm) from the crop rotation experiment. Soils from the cereal-fallow rotation contained the lowest levels of all the microbial properties measured, whereas the cereal-vetch rotation supported a higher level of microbial biomass and activity (Figure 4). As shown here, increased levels of microbial biomass are associated with improved crop yields.
Figure 4. Averaged over 3 seasons, soil microbial biomass carbon was 48% higher in the legume systems compared with the cereal-fallow systems. Cotton lint yield was 16% higher in the legume systems.
Soil from the oats-vetch rotation not only contained more N in the microbial biomass (34 kg/ha more than the oats-fallow rotation) but also exhibited highest level of N mineralization potential (71% more than the oats-fallow rotation – Table 3). The soil from the continuous vetch-cotton-vetch rotation exhibited intermediate levels of microbial activity and N mineralization potential.
Microbial quotient (MQ) is considered a more useful measure to indicate changing soil microbial processes across management practices as it normalizes the microbial biomass values across different organic matter contents. Data in Table 3 indicate higher MQ value for the soil from the oats-vetch system. In the majority of Australian agricultural soils, the biota is constrained by the lack of biologically-available organic matter. Hence, the cropping systems that provide more carbon inputs (above-ground and below-ground) generally support higher levels of microbial biomass and microbial activity.
Table 3. Rates of soil N mineralisation, microbial respiration and the proportion of soil carbon contained in the soil biomass (Microbial quotient).
|
Cropping system
|
Potential N mineralisation
(µg N / g soil / day)
|
Microbial respiration
(µg CO2/g /day)
|
Microbial quotient
(MB-C/Organic C)
(%)
|
|
oats-fallow-cotton
|
0.42
|
8.65
|
3.31
|
|
oats-vetch-cotton
|
0.72
|
13.20
|
4.63
|
|
vetch-cotton-vetch-cotton
|
0.56
|
10.26
|
4.11
|
Nitrogen from fertilizer sources is prone to losses such as leaching, volatilization and denitrification whereas the N from the microbial biomass is less prone to such losses. Since N availability is one of the major regulating factors of crop yield, especially for irrigated cotton crops, it is not surprising to see a significant relationship between microbial biomass-N value and lint yield.
The oats-vetch rotation not only supplied a larger total amount of C but also crop residues of higher quality (i.e. C:N ratio) and therefore promote a more active and diverse microbial community. The three crops (cotton, oats and vetch) harbor different types of microbial communities, due largely to the different compounds exuded from their root systems that produce distinct rhizosphere effects. In dryland grain and pasture systems, it has been shown that diverse plant types generally support a greater functional diversity in soil biota communities.
Conclusion
There has been no single soil characteristic that we have measured that can explain the cotton yield benefits of growing rotation crops, particularly with legumes. We have observed improved soil structure, nutrient availability and uptake, a more active and larger soil microbial biomass which combine to improve soil health, and enable future crops to be more productive and use inputs more efficiently.
References
Rochester I.J., Peoples M.B., Hulugalle N.R., Gault R.R. and Constable G.A. 2001. Using legumes to enhance nitrogen fertility and improve soil condition in cotton cropping systems. Field Crops Research 70: 27-41.
Rochester I.J. and Peoples M.B. 2005. Growing vetches (Vicia villosa Roth) in irrigated cotton systems: inputs of fixed N, N fertiliser savings and cotton productivity. Plant and Soil 271: 251-264.
Sparling G.P. 1997. Soil microbial biomass, activity and nutrient cycling as indicators of soil health. In: Biological indicators of soil health. C.E. Pankhurst, B.M. Doube and V.V.S.R. Gupta (eds.). pp. 97-120. CAB International, UK.
Contact details
Ian Rochester1
CSIRO Plant Industry, Cotton Catchment Communities CRC
Australian Cotton Research Institute, LB 59 Narrabri
Ph: 02 6799 1520
Email ian.rochester@csiro.au
Was this page helpful?
YOUR FEEDBACK