SOIL ADDITIVES TO STIMULATE N FIXING AND P AVAILABILITY
| Date: 16 Sep 2009
Preamble - the legal problem presented by speaking about products
One of the first constraints to a scientist in presenting a paper like this, is the risk that the statements made, may be viewed by those with a commercial interest in selling products to farmers, as so damaging (or so wrong) that they need to respond by taking legal action against the scientist. Even the threat of legal action may be so daunting that the scientist or the employer may prefer to reman silent. One of the most celebrated examples of this is the Maxicrop case, in which the Bell-Booth Group sued the New Zealand Ministry of Agriculture Fisheries (MAF) and Dr Doug Edmeades personally for damages (initially $5.5 million, later amended to $11.5 million). This story is wonderfully presented in Edmeades’ book “Science Friction. The Maxicrop case and the aftermath” (Edmeades, D.C. 2000. ISBN 0 473 06886 9, Published by Fertiliser Information Services Ltd., P.O. Box 9147, Hamilton, New Zealand), and the underlying science published in the Australian Journal of Agricultural Research – Edmeades, D.C. 2002. The effects of liquid fertilisers derived from natural products on crop, pasture, and animal production: a review. Aust J Agric Res 53:965-976. Here is a potted history of the story.
Maxicrop is a concentrated seaweed extract, which was promoted as a fertiliser, providing nutrients and plant hormones. The recommended application rates, highly diluted, meant that it was considerably cheaper than conventional fertilisers. As with farmers everywhere, New Zealand farmers in the mid-1980s were subject to economic pressures, and with fertilisers as a major cost a cheaper alternative was welcomed. After extensively reviewing the world literature on non-traditional fertilisers, analysing Maxicrop, and undertaking field trials with it, Dr. Edmeades came to the conclusion that, used as directed, the product could not possibly provide the claimed benefits. In April 1985 Dr. Edmeades appeared on the TVNZ program ‘Fair Go’ with Mark Bell-Booth and David Bellamy in which he presented his case against Maxicrop. It was this program which provided the basis for the subsequent legal action.
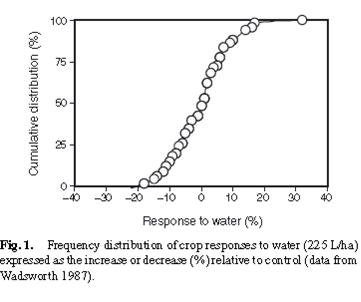
One aspect of the company’s case was the claim that Maxicrop did work in some situations (increasing crop yield), and while they could not accurately predict which conditions it would work under, there was nevertheless evidence that the product did work. The real difficulty for Doug Edmeades and MAF was to explain to people trained in law, rather than natural sciences, that natural variability (in statistical terms – error) would result in Maxicrop occasionally producing a yield greater that the control. One of the key pieces of information that helped the lawyers grasp this idea was a set of data showing the response of crops to an application of water (Figure 1, from Edmeades 2002 ).
Figure 1 is a frequency distribution relationship; it shows how frequently a particular response to the application of a small amount of water was obtained. From Figure 1, you can see that occasionally application of water reduced crop growth (by almost 20% in a very small number of instances). Conversely, water also increased yield in some instances. The key aspect of this set of data is that the data points are centred around zero (the mean is actually -0.6% and confidence interval 2.3%). In fact, application of water had no effect on yield, the range of results obtained is consistent with variability normally associated with this type of experiment.
Using this basic understanding of variability, it was apparent to the court that Maxicrop did not work, and that the occasions when it appeared to increase yield were simply random variation (experimental error). Frequency distributions for Maxicrop and several other similar liquid fertilisers are presented below overlaid with the response to water in each of the trials (Figure 2 from Edmeades 2002). Overall a great demonstration that materials of this type (low nutrient concentration, and compounds intended to act as plant growth stimulants) are not effective fertilisers. 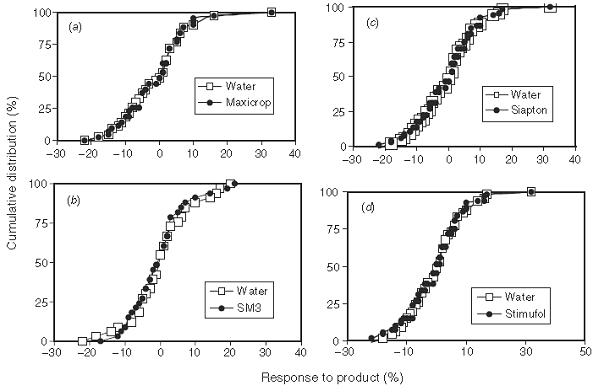
Figure 2. Frequency distributions for Maxicrop (seaweed), Siapton (animal offal extract), SM3 (seaweed), and Stimufol (vegetable)
Ultimately, the judgement mostly went against the plaintiffs except in one regard. In relation to the claim of negligence the judge, Justice Ellis, stated, “MAF is in the most general way under a duty to act fairly to all citizens. This involves balancing competing interests. The present case is a good example. MAF must in my view balance its primary obligations and duties to the pastoral and agricultural industries and to the vendors of products consumed by such. In general terms I consider that where an agency such as MAF intends to condemn a product it must give the seller an adequate and fair opportunity to consider such publicity beforehand and make its responses before the damage is done.” Consequently his Honour found that MAF had breached this duty of care and had acted negligently, awarding the Bell-Booth Group $25,000.
The company was not satisfied with this tiny victory and appealed to the Court of Appeal, which provided no encouragement and overturned the negligence verdict, observing inter alia that, “Some of the arguments for the company go close to asserting that a manufacturer has a right to sell worthless goods as long as he honestly believes that they are some use. We would see that as putting it over-simply. Those who reasonably believe that the goods are worthless must have an equal right to say so.” Despite the clear defeat of the plaintiff, who had initiated the case, the view was widely expressed that the powers of government had been used to crush a small struggling entrepreneur.
Despite the eventual court decision, the case cost Doug Edmeades at least 18 months of his professional career; it was undoubtedly also a most harrowing experience. Further, as explained by Dr Edmeades in ‘Science Friction’ MAF were subsequently very hesitant to allow their scientists to publish results that did not support a commercial product. “How does this help farmers?” we might well ask. We have no desire to follow in Doug’s footsteps, so we are setting some rigorous rules in what we intend to say and how we can justify these statements. Throughout this paper, we will stick to reporting to you the “science” (Doug Edmeades also did this, but it still was not enough to keep him safe). We will not refer to specific products. However, we do want to provide you with something useful, so we will attempt to provide you with a way to think about products which are intended to improve plant growth through their influence on soil biology. We also make some suggestions of how to go about testing these products yourself.
The nature of Science and of Faith – we will stick to science.
Usually "faith" refers to beliefs that are accepted without empirical [observed] evidence. Most religions have tenets of faith; but there are many other belief systems not supported by empirical evidence. Science differs from these belief systems because it is the nature of science to test and retest explanations against the natural world. Thus, scientific explanations are likely to be built on, and modified with, new information and new ways of looking at old information. This is quite different from most religious beliefs. Importantly, as scientists, we must always be ready to throw out our most cherished ideas – even if we have worked on them for a lifetime.
The basic nature of science is to form a hypothesis which explains an observation; this hypothesis is then tested. If through repeated testing the hypothesis is shown to account for the observations, then it is regarded as a theory. In scientific terms, "theory" does not mean "guess" or "hunch" as it does in everyday usage. Scientific theories are explanations of natural phenomena built up logically from testable observations and hypotheses. Scientists generally use the term "fact" to mean something that has been tested or observed so many times that there is no longer a compelling reason to keep testing or looking for examples.
When we are sure of a “fact” (and often before we are sure), scientists often want to know “why”, and this is very important because we must find the underlying principles behind an observation. This has two benefits. First, it allows us to more clearly decide when the same treatment is likely to be beneficial in another situation (or, as important, when not). Second, it allows us to confirm our finding by testing it against known principles (even though we recognise that these principles can be overturned). As eloquently stated by Richard Feynman “The whole question of imagination in science is often misunderstood by people in other disciplines. They overlook the fact that whatever we are allowed to imagine in science must be consistent with everything else we know.” (from Genius by James Gleick). Finally, it is important that information be published in peer-reviewed, scientific journals. As stated by Horace Judson: “Formal publication – formal invitation to criticism” (The Search for Solutions). Of course, peer review has its drawbacks – when powerful opponents to new ideas prevent publication.
Therefore, "belief" is really not an appropriate term to use in science, because testing is such an important part of this way of knowing. If there is a component of faith to science, it is the assumption that the universe operates according to regularities. . . . This "faith" is very different from religious faith. (From - National Academy of Sciences Working Group on Teaching Evolution. Teaching about Evolution and the Nature of Science. Washington, DC: National Academy Press, 1998.)
In contrast, faith is something one "believes in". It serves a major evolutionary purpose and has been an essential part of human nature since time immemorial. When shared by members of a group, faith strongly supports that group's internal cohesion. It strengthens the group's capacity to cope with the challenges of a hostile environment. It adds to the group's capacity to compete successfully with other groups animated by different faiths. But there is a dark side to the ‘in group’ – whether religious or not – by definition, there is an ‘out-group’. A basis for hostility – often extreme – if we are not careful. All religion is based on faith, but not all faith needs to be religious, at least in the sense of requiring adherence to a recognised religious persuasion. (From - Carl Croon – Progressive Humanism)
Let’s briefly look at examples that are readily accepted as sitting near the opposing ends of this Science – Faith spectrum. The Rhizobium – Legume symbiosis is a good example of science. In this unique association between organisms, the plant provides a source of energy and an ecological niche for the bacterium, which in return synthesises ammonia for the host plant. Despite millions of years of evolution, higher plants have not developed a N2-fixation system. At the global scale, the Rhizobium-legume symbiosis provides a quantity of fixed nitrogen (N) comparable to that produced by the entire chemical fertiliser industry, and thus plays a major ecological and economic role (Table 1). The symbiosis has been the subject of a great deal of scientific investigation, and we now understand it at a genetic, biochemical and ecological level. Furthermore, we understand it well enough that the system can be, and is, effectively manipulated in farmers’ fields all over the world.
Table 1. The major annual terrestrial N inputs. Fixation by legume crops is subdivided to show the importance of soybean. (Simil 1997;Herridge et al 2008)
|
Source
|
|
Nitrogen fixed (Tg)
|
|
|
Natural biological N fixation
|
|
|
90 - 140
|
|
Lightning
|
|
|
<10
|
|
Symbiotic N fixation by crop legumes
|
|
|
22
|
|
|
Soybean
|
16
|
|
|
|
Peanut
|
2
|
|
|
|
chickpea
|
0.6
|
|
|
Pasture legumes
|
|
|
12-25
|
|
Fertiliser
|
|
|
160
|
Symbiotic N2-fixation
Symbiotic N2-fixation must be one of the most wonderful processes in biology, and it’s the association between Rhizobium (a bacterium) and legume (a higher plant) that most impacts on agriculture. This is a true symbiosis – each organism gains an advantage – the Rhizobium with energy and the legume with fixed N. It’s worth considering the great effort that has been made to ensure that our efforts to manipulate the symbiosis work. Let’s stet through the important parts of the process.
The scientist must first identify strains of Rhizobium that can more effectively fix N in the field situation. This sounds a contradiction to what was stated earlier, but we can ensure improved conditions such that the bacteria grow better – i.e. to their advantage. The scientist selects nodules from the roots of a legume growing well apparently because of N2-fixation (Plate 1), crushes them, and grows the released bacteria in an artificial medium. Purified isolates (strains) are then selected for a number of important characteristics which include ability to infect, nodulate and fix N2 with the a range of legume cultivars in use, genetic stability of the strain over time in culture, ability to survive in the inoculum formulation (ie shelf life) and after inoculation on the seed before sowing and after planting in dry soil. A further selection criteria is then the ability of the inoculum strain to compete with other rhizobial strains in the soil, which are less effective in fixing N2, to multiply in the rhizosphere and form the nodules. Lastly the strain is selected for its ability to survive in the soil between seasons. The latter is more important for pasture legumes. Once a superior rhizobial strain has been identified and selected, the inoculum production has to be done properly.
Rhizobia grow in culture relatively slowly (3 to 5 days to come to maximum numbers in a fermentor) compared to many bacteria and fungi that are quite ubiquitous, and hence potential contaminants – the pseudomonas, bacillus, penicillin species that grow up overnight. Thus it is necessary to have very good quality control in the production system to avoid producing contaminant organisms rather than Rhizobia. Australia has the best system in the world for this process. This is mainly because of the very close relationship between the researchers and the State Departments concerned with Agriculture. A key aspect was the establishment of an inoculum quality testing system (undertaken by the Australian Legume Inoculant Research Unit) that is a unique voluntary testing system that all manufacturers use, and they abide by the very high inoculum quality standards required for sale. This partnership has resulted in the evolution of inoculants mainly as a sterile (gamma irradiated) peat inoculated with the broth from the fermentor. Peat is the best material found for multiplying and preserving rhizobia both in the packet and on the seed. However, new forms of inoculant are now being tested in Australia, a paste of rhizobia or a concentrated broth that is mixed with water and used as liquid in furrow inoculant at sowing, or a granular inoculant that is run into the furrow with the seed. It is the responsibility of the agronomist and the farmer to check that these new inoculation methods work well to produce nodules that are near the crown of the plant and are effective (pink when cut open but more of this later).
Finally, it is the turn of the farmer. The good farmer ensures that the full recipe of packet instructions is followed: not allowing the peat inoculum to heat up, or dry out, or be exposed to the sun; adding a methyl cellulose sticker to the peat inoculum when it is mixed with water and then the seed (sometimes a concrete mixer is used for this); drying the inoculated seed in the shade, to minimise heat stress and effects of UV; then planting with minimum delay. The overall aim is to ensure that as many as possible viable bacteria are on the seed in the planting furrow. Alternately some farmers are now mixing the peat directly with seed as it is augured into the seed boxes on the planter - if a satisfactory inoculation is to be obtained the mixing has to be thorough and the inoculant must stay distributed throughout the seed in the box.  Plate 1 (left). Nodules on a soybean root, with one cut in half to reveal the cortex (thin, pale outer structure) and the pink leghaemoglobin that indicates bound oxygen and properly functioning N2-fixation. Cut nodules that are brown or green because the haemoglobin has broken down are not N2-fixing.
Plate 1 (left). Nodules on a soybean root, with one cut in half to reveal the cortex (thin, pale outer structure) and the pink leghaemoglobin that indicates bound oxygen and properly functioning N2-fixation. Cut nodules that are brown or green because the haemoglobin has broken down are not N2-fixing. 
Plate 2 (right). Light micrograph of a cowpea root stained with crystal violet showing the mucilage excreted mostly from the root cap, and which is an important source of energy for rhizosphere microorganisms including rhizobia.
The final process in this wonderful saga is now ready to take place. The seed will imbibe water, and a radical (root initial) and plumule (shoot initial) will break through the seed coat. The radical will be ‘contaminated’ with the bacteria on the seed coat, and they will get first access to the carbohydrate-rich mucilage excreted from the root (Plate 2); they will grow and divide, so increasing their competitive advantage over other rhizobial strains and other microorganisms that are in the soil and grow in the rhizosphere in the soil. The radical will grow and become the primary root which has a zone of cell division, cell elongation, and a zone where root hairs develop. In a legume like soybean, the root hairs are where the bacterium starts the infection process by causing the root hair to curl with the subsequent development of an infection thread. The bacteria multiply within each thread, which invades the root cortical cells inducing them to divide in response to the infection. The rhizobia are released from the thread and become encapsulated within a plant cell membrane, continue to divide, and at last, the N fixing genes are switched on and nitrogenase enzyme produced. This requires an adequate supply of iron, an essential part of the enzyme and of the leghaemoglobin that gives the sliced nodule its pink colour. All this involves signals between the legume and the rhizobia (both of which must be genetically compatible), and the scene is set for the nodule to start developing. This is where the trading takes place – the plant supplies energy in the form of carbohydrates to the rhizobia; in return, they fix N2 into a form that can be used by the plant.
For the symbiosis to be successful there is a major balancing act required: aerobic (oxygen (O2) consuming) respiration is much more efficient than anaerobic respiration but the nitrogenase must be protected from O2. The magnitude of this protection is astounding – and the balancing act, even more so. The O2 concentration in the soil solution is ca. 360 µM, and this might halve inside the root due to respiration. But this concentration is still far too high and would denature the nitrogenase. It’s in the nodule cortex (Plate 3) that the initial barrier is set up – by all intercellular spaces filled with water and suberised (corky) cells, for example – and this barrier ensures that the O2 concentration in the nodule is low. Even more extraordinary, there is a compound in the bacteria filled cells of the nodule, called leghaemoglobin (it’s very similar to the myoglobin in our muscles) which binds the O2 molecules entering the interior of the nodule, such that the free O2 concentration is about 10 nM. This is a little less than forty thousand times lower than that just outside the nodule, less than 1 mm away. It’s the leghaemoglobin that gives an effective nodule its pink colour (Plate 1). This concentration of residual O2 is enough to diffuse through to the bacteria to support aerobic respiration – but not enough to poison nitrogenase. 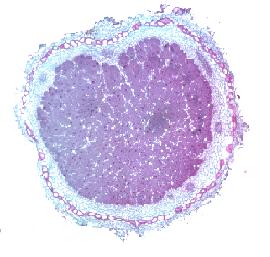 By all standards and without human intervention, this is an astoundingly complex process. It works! – and for over a century, we have been able to make it work to our advantage. No wonder we look to other bacteria and fungi in like manner.
By all standards and without human intervention, this is an astoundingly complex process. It works! – and for over a century, we have been able to make it work to our advantage. No wonder we look to other bacteria and fungi in like manner.
Plate 3. Cross section through a legume nodule. showing the purple stained lignified cells which surround the nodule and limit oxygen entry, and the vascular bundles which supply the bacteria with their substantial energy requirement. The core area of the nodule is where the bacteria fix N2. Within this core area, the plant cells (stained mauve) are packed with the N2-fixing bacteria and leghaemoglobin
At the other extreme, homeopathy is viewed by scientists as representing an example of faith. A central thesis of homeopathy is that an ill person can be treated using a substance that can produce, in a healthy person, symptoms similar to those of the illness. Practitioners select treatments based on consultation that explores the physical and psychological state of the patient (not a bad idea!), both of which are considered important to selecting the remedy. According to Hahnemann, one of the key figures in the development of the approach, serial dilution, with shaking between each dilution, removes the toxic effects of the substance, while the essential qualities are retained by the diluent (water, sugar, or alcohol). Claims to the efficacy of homeopathic treatment beyond the placebo effect are unsupported by the collective weight of scientific and clinical evidence. Common homeopathic preparations are often indistinguishable from the pure diluent because the purported medicinal compound is diluted beyond the point where there is any likelihood that molecules from the original solution are present in the final product; the claim that these treatments still have any pharmacological effect is thus scientifically implausible and violates fundamental principles of science. Critics also object that the number of high-quality studies that support homeopathy is small, the conclusions are not definitive, and duplication of the results, a key test of scientific validity, has proven problematic at best. The lack of convincing scientific evidence supporting its efficacy and its use of remedies without active ingredients have caused homeopathy to be regarded as pseudoscience or quackery (From the reference most loved by university lecturers – Wikipedia). Also, in some instances, a belief in the benefit of a treatment is considered a prerequisite for its efficacy – this precludes scientific testing.
The dilutions advocated in homeopathy are extreme. A 1060 dilution was advocated by Hahnemann for most purposes. Some trivial calculations put this dilution in context – if you used a medicine diluted to 1060, you would need to give two billion doses per second, to six billion people (the world’s population), for 4 billion years, to deliver a single molecule of the original material to any patient. One third of a drop of some original substance diluted into all the water on earth would produce a remedy with a concentration of about 1026 (once again I am trusting Wikipedia for this value).
The difficult middle ground
The extremes are easy – we can readily accept or reject ideas (or products) for which there is clear evidence and understanding on which to base our decision. Our decision-making task is much more difficult when there has been only limited investigation, and hence there is little information, or when the results of investigations are inconsistent.
For the two examples we will develop here – the fixation of atmospheric nitrogen gas (N2) and P-solubilisation by free living organisms – there is clear evidence that these processes do occur. The question we will attempt to address is: Can these processes be manipulated and enhanced? From the perspective of a farmer, the question would be: Are products which claim to do this, worth the investment?
But first some thoughts about the nature of the plant-microbe- soil interactions Plants can grow without microbes on the roots, but it is extremely difficult to do so and it took the NASA space program to stringently attempt to do this with a modicum of success. Most seeds have embedded bacteria and it is very difficult to “sterilise” them. Likewise a large volume of soil is very difficult to sterilise, and when plants are grown in soil almost impossible to keep microbes from contaminating the plant growth system. Enclosed sterile plant growth chamber systems are required and it is very difficult to provide enough light to saturate the plants requirement for maximum photosynthesis. Hence it is very difficult to compare growth of plants with and without microbes. Plants growing with microbes, is the natural condition.
Plant roots are surrounded by a mucilaginous layer, the “mucigel” mainly exuded from root tip (Plate 2). The space immediately surrounding the root, where microbes grow in greater numbers than in the bulk soil, is known as the rhizosphere which usually extends about 1-2 mm from the root. The plant exudate contains a wide range of amino acids, sugars, organic acids and vitamins. Some bacteria are selectively stimulated to multiply by this substrate. The amount of carbon in the photosynthate exuded into the rhizosphere can be as much as 25-30% of the total amount fixed by photosynthesis.
So much of the plants energy is invested in rhizosphere functions. One of these is to develop a population of microbes, both bacteria and fungi, which protects the plant from infection by pathogens and plays a role in plant nutrition and plant growth stimulation through production of plant hormones. There is a homeostatic process operating in the rhizosphere so that the numbers of a particular organism reach an equilibrium level. The microbes respond to plant signals which affect their gene expression, and also through a process known as quorum sensing, where microbes limit their own population development once a certain level has been reached. Most of the organisms living in the rhizosphere and the bulk soil cannot at present be grown in culture medium. The genetic diversity of the soil microbial population can be affected by the farming system, soil type and plants grown. So we have a very complex system which, as we shall show with our P uptake example, has feedback interactions also. There is a lot we do not know about the plant microbe interactions in the soil. So the way to see if we can manipulate the system to be economically beneficial for sustainable agricultural production is to undertake very well designed empirical experiments properly replicated over time and environment, soil type and farming system.
In the bulk soil and rhizosphere the microbes are responsible for mineralising (breaking down) organic matter, thereby releasing nutrients for plant uptake and growth. The benefits accruing for plant growth from richly organic soils is directly the result of microbial activity. However, all of these benefits are from microbes existing naturally at sufficient populations in soils to undertake this process of mineralisation, growth stimulation, pathogen control, etc. For processes like mineralisation, there is invariably no need to add any more micro-organisms to the soil. In the following sections we explore the question, of whether adding additional organisms is ever effective. We develop this question in our two examples of plant microbe interactions – it’s for you to judge if we have managed to answer it.
Free-Living Nitrogen Fixers
Nitrogen fixation. Prior to the development of the Harber-Bosch process (Smil 1999) for the industrial fixation of N, biological N2-fixation was the primary means by which atmospheric N2 gas was converted to ammonium, and most life on earth ultimately depended on this process. At least 90 genera of microorganisms produce the enzyme nitrogenase, and are hence capable of fixing atmospheric N2. These microbes are free-living organisms, or in a loose association with lichens, mosses, the fern Azolla, cycads and some higher plants (associative N2 fixation), and in complex symbioses with a host plant (symbiotic N2 fixation). The most important N2-fixing process in agriculture is the symbiotic association between crop and fodder legumes and rhizobia. At a global scale symbiotic N2-fixation by crop legumes is estimated at 21 Tg (one teragram equals 1 million tons), with pasture legumes fixing another 12 to 25 Tg. Non-symbiotic N2-fixation is less important, and is estimated to be responsible for the fixation of about 20 Tg of N2, of which <4 is thought to occur on non-legume crop fields (cereals) (Herridge et al 2008).
Free-living nitrogen fixers. Now let’s consider how much N free living organisms may fix in a typical agricultural field in this part of the world. As a starting point, we need to consider why an organism would fix N – “Because that’s what they do.”, is simply not a good enough answer. Unfortunately, we are often told to regard soil organisms as a wonderful community of life in the soil helping us to grow a healthy crop – a viewpoint redolent in environmentally acceptable overtones. What utter nonsense! Soil organisms may do a great deal which results in the production of a healthy crop, but whatever they do, you should consider that they are doing it for their own good, not yours. Not for my good, not for yours, not for any other reason than their own survival.
Soil organisms fix atmospheric N2 gas in reasonable amounts only when mineral N is limiting in the soil (and in the organic matter they are living on). This preference for soil mineral N, over fixation of atmospheric N, is driven by the high energy cost of N2-fixation. The energy cost of N2-fixation by white clover (Trifolium repens) has been estimated to be 25% of all of the carbon fixed in photosysthesis per day. This cost should be compared to the energy cost of taking up N as nitrate or ammonium from the soil, estimated at 4 to 13% of the carbon fixed by photosynthesis when the N supply is good (Lambers et al 2008). This energy cost of taking up soil N increases as fixed N becomes limiting (i.e. the plant needs to invest more in translocating sugars to the roots or in growing a bigger root system, etc), to values comparable to, or exceeding, the cost of N2 -fixation (Lambers et al 2008). The efficiency of N2-fixation by free-living organisms is about one tenth of that in legume symbiosis (Silvester and Musgrave 1991). Given this energy cost of N2-fixation, an organism would only spend energy in this way if the supply of N in the soil was limiting – and, of course, it would need to have a good food supply in order to have the energy to spend (eg root exudates or crop residues - or sunlight for its own photosynthesis in the case of cyanobacteria).
Nitrogen fixation also requires a low oxygen environment for the enzyme to work; nitrogenase activity is very low in an oxygen-rich environment. In the legume-rhizobia symbiotic relationship, the nodule protects the bacteria from exposure to oxygen (as we discussed earlier). For the free-living N2 fixers, some require anaerobic (oxygen free) conditions, others such as Azospirillum are capable of fixing N2 in low oxygen environments (microaerophillic). Some bacteria produce gum to provide an oxygen barrier. Some aerobic bacteria such as Azotobacter have very high respiration rates (10 to 50 times greater than other organisms) and can in this way lower their intracellular oxygen to a level sufficiently low for N2-fixation to occur. Protection of the nitrogenase from oxygen typically comes at a high energy cost, and this limits bacterial growth, and hence limits N2-fixation. Hence non-symbiotic fixation is much lower in aerated soils than in wetter, less aerobic soils (Einsele et al 1996)
Thus, we can define some conditions required for N2-fixation by free living organisms to occur at a reasonable rate – high energy (carbon) supply, low mineral N, low oxygen supply, and a moist soil, the more the better. The conditions of limiting N availability and good carbon supply, exist for much of the year in forests and grasslands; the established root systems of the plants scavenge any free mineral N in the soil, and there is a continuous input of organic matter through leaf drop and root turnover. Estimates of N2-fixation in forests and in grasslands vary widely, and are typically in the order of 0.01 to 5 kg/ha/year (Son 2001) but can be as much as 39 kg/ha/yr (see review by Dart 1986). In winter cereal cropping systems, the conditions of low mineral N and high energy supply (high carbon supply) are met at the start of the summer fallow. However, at this time the soil is dry, limiting biological activity, and exposing potential free-living N2-fixing organisms to a high oxygen environment. During the fallow period, the soil water store builds, and the extent of oxygen exposure will decrease – but the mineral N pool in the soil also builds up during the fallow, and this supply of N will be far more energetically attractive to organisms than obtaining N through fixation. Conditions of low oxygen and N, and a good energy supply, can also exist near the roots of plants (the rhizosphere). Oxygen levels are lowered by plant root respiration, N concentration is lowered by plant uptake of mineral N, and root exudates supply an energy source.
Estimates of N2-fixation in cereal fields range from <5 kg/ha/y (e.g. Lamb et al 1987) to 30 kg/ha/y (Dart 1986). In Australia, there have been very few field studies, because of the methodological difficulties. The conversion factor to convert between the experimental measurement of nitrogenase activity, acetylene reduction by nitrogenase, and the actual level of N2-fixation which is occurring, is difficult to quantify. Nitrogen mass balance studies often do not measure all the inputs and potential losses, or examine the soil profile at depth, so they are an indication only. In an assessment of the non-symbiotic N2-fixation in the winter dominant rainfall environment of South Australia, Gupta et al (2006) estimated 20 kg N/ha/y was fixed in continuous wheat cultivation. Subsequently, Unkovich and Baldock (2008) reviewed existing literature and suggested that a figure of <10 kg/ha/y may be more appropriate. In many respects this discussion has little relevance to a farmer. Irrespective of what the number is, this has been occurring in the field without intervention, and the appropriate fertiliser application rate for the system (determined by trial or experience) already includes this input. In other words, a sensible farmer on learning that organisms in the soil were fixing 10 kg N /ha/y would not respond by reducing fertiliser N input by that amount.
Effectiveness of seed inoculation with free-living organisms. Finally, let’s consider if there is a need for inoculation of the crop with free-living N2-fixing organisms. This would be necessary if the soil did not contain organisms capable of fixing N2, or if those that did exist in the soil were not particularly effective in this role. There has been a reasonable amount of work on the first aspect – whether or not there are N2-fixing organisms in Australian soils. Gupta et al (2006) reviewed the work in this area and reported that non-symbiotic N2-fixing bacteria were present in all soils sampled regardless of soil type or land use (Figure 3). Soil type did have an influence on the population of N2 fixers, with clay soils having the largest populations, and sandy soils the lowest. However, land use had a greater effect on the population (103 to 105 MPN/g –most probable number per gram), with land used for cereal production having higher populations than land used for legume production – the effect you would expect considering our earlier discussion of the energy cost of fixing N2 relative to using mineral N present in the soil. Given these substantial existing populations in the bulk soil, inoculation of the seed is unlikely to have a large impact on the size of the population of N2 fixers.
So we come to the second aspect, whether or not we can improve N2-fixation by inoculation with organisms that develop a larger rhizosphere population with greater N2-fixation ability. This is a tempting prospect, and it certainly has been the experience with the legume/rhizobia symbiosis, where inoculation with a competent strain of rhizobium can improve crop yields, even in fields where nodulation would occur without inoculation. Various scientists have proposed that analogous (though clearly much smaller gains) could be achieved for non-symbiotic N2-fixation (Kennedy and Islam 2001, Triplett 2007). To date, research demonstrating that worthwhile gains are achievable as a result of seed inoculation, has not been published. While the zone around plant roots does provide a suitable environment for N2-fixation (low oxygen, low N, and good energy supply), our understanding of the high energy cost of the N2-fixation process, and the relatively low efficiency of free-living N2 fixers, would lead us to expect that the amount of N2 fixed in the root zone of a cereal crop will always be modest. 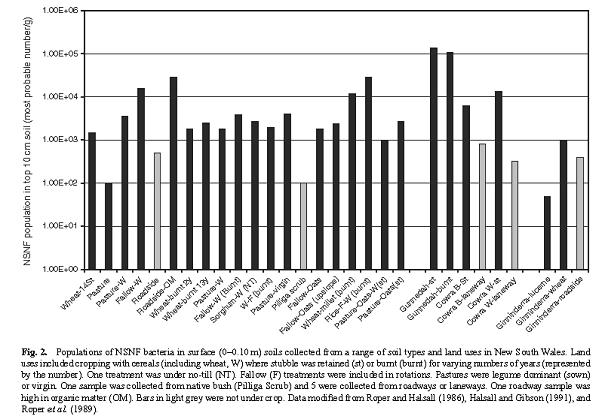
Figure 3. Populations of free living N2-fixing bacteria in surface soil samples from a range of soil types and cropping systems (from Gupta et al 2006).
Phosphorus solubilising organisms
After N, phosphorus (P) is the most commonly limiting nutrient in soils around the world. Soils typically have a reasonably large store of P, many soils contain a total P store sufficient for 100 years of cropping, but the problem is that most of this P is not in a form which is available for plant uptake. Plants take up P directly from soil solution as orthophosphate (HPO42- and H2PO4-). As the soil solution P is depleted, other pools of P that are held on the solid phase of the soil, but are in equilibrium with the soil solution, will be released into solution. For example, P that is adsorbed to soil minerals desorbs, thus buffering the P in the soil solution. Another pool of P in the soil is organic-P which, like N in the organic matter, needs to be mineralized in order to be available for plant uptake. Typically, 20 to 70% of the total soil P is in this organic pool in mineral soils (values for organic soils and peats are much higher, of course). Many plants, including wheat, can achieve the release of part of this organic-P through the release and action of the enzymes called phosphatases. Production of phosphatase is enhanced by low P conditions. The phosphatase enzymes achieve release of orthophosphate from nucleic acids, phospholipids, glucose 1-phosphate, and glycerophosphate, all of which are present in soil, and were derived from cell cytoplasm. However, they account for only a minor part of the soil organic-P and generally are not present in soil in sufficient quantities to supply an actively growing plant’s needs. Most of the remaining organic-P cannot be readily accessed by the roots of most plants, but release by the plant of root exudates acts as an energy source for organisms which are able to produce enzymes capable of releasing this organic P. The concentration of organic P near the roots of wheat can decrease dramatically (by 86% in a study by Tarafdar and Jungk 1987). So organisms which mobilise organic-P are clearly important. In this paper we will concentrate on organisms which are capable of mobilising P from inorganic forms.
Plant P solubilisation
Some plants can themselves solubilise inorganic P from the soil solid phase in order to make it available for uptake. We will start by considering this simpler system (soil?_plant), before considering the more complex system of release by a micro-organism (MO) and eventual capture by the plant (soil?_MO?_plant). Some plants that are adapted to low P soils, for example lupin (Lupinus albus) excrete acidifying or chelating compounds (e.g. citric and malic acids – Figure 4) (chelation is binding or complexation of a metal through a bi- or multi-dentate bond). Most plants (including wheat and maize) do not appear to do this. Acidification can solubilise P in alkaline soils, but this strategy is not effective in acid soils. The chelating organic acids are able to compete with P for adsorption sites, releasing P into solution. They also complex metal cations (Ca, Al, Fe) resulting in the dissolution of soil minerals – some of which may contain P (e.g. calcium-phosphate minerals such as apatite, aluminium-phosphate minerals such as variscite, and iron-phosphate minerals such as strengite). Members of the plant family Proteaceae are particularly effective at producing and excreting organic acids into the root zone. Plants of this family (and a number of other families) are able to form cluster roots (Plate 4). This structure permits the effect of organic acid release to be concentrated in a limited volume of soil to maximize its effectiveness. We can regard these plants as mining P, by forcing its release from the solid phase. In contrast, the mycorrhizal associations of many crop plants could be considered as scavangers, picking up whatever free P (P in the soil solution) they can find. Mycorrhizal associations are more effective in soils where the soil solution P concentration is somewhat higher than that in soils where Proteaceae are abundant (Lambers et al 2008).
It’s worth considering the energy (carbon) cost to the plant of obtaining P by different strategies. In soils with a reasonable P status, the roots and root hairs are sufficient to obtain sufficient P. As P becomes more limiting, the plant will invest more of the carbon it fixes through photosynthesis, and this can be seen in an increased root to shoot ratio (more roots and fewer shoots). In still lower P environments, mycorrhizal associations are beneficial to the plant – it costs less energy to support a network of fungal hyphae than it does to build a system of roots and root hairs. For mycorrhizal plants, 4 to 20% of carbon fixed in photosynthesis is used by the mycorrhiza. It is interesting to note that the formation of the plant-mycorrhizal symbiosis is affected by P supply; plants do not form an association in high P soils, as feeding the fungus would represent an unnecessary expenditure in this situation. Finally, the production of cluster roots and release of organic acids is extremely energy expensive; the plants strategy is to acquire P at any cost (Lynch and Ho 2005). It is no surprise that plants which do this are slow growing. Energy cost is clearly critical when we are considering a crop, as any additional investment in obtaining nutrients can reduce yield. 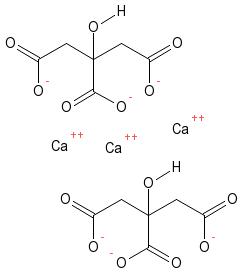 Figure 4. The citrate molecule has three negative charges, and is hence capable of forming a multi-dentate bond to Ca2+ (or other cations such as Fe3+ and Al3+). The calcium-citrate is soluble, so the formation of this complex will result in the dissolution of minerals such as apatite (the calcium phosphate in rock phosphate). Note that each bend in the drawing of the citrate molecule is a carbon atom (six in total) giving an indication of the energy cost to the plant of producing and releasing this molecule.
Figure 4. The citrate molecule has three negative charges, and is hence capable of forming a multi-dentate bond to Ca2+ (or other cations such as Fe3+ and Al3+). The calcium-citrate is soluble, so the formation of this complex will result in the dissolution of minerals such as apatite (the calcium phosphate in rock phosphate). Note that each bend in the drawing of the citrate molecule is a carbon atom (six in total) giving an indication of the energy cost to the plant of producing and releasing this molecule.

Plate 4. Cluster roots grown in solution culture (left) and in soil (right). (from Lambers et al 2008).
Free living solubilisers.
Micro-organisms capable of solubilising P are ubiquitous in soils, with 1 to 50% of the total bacterial population, and 0.1 to 0.5% of the total fungal population capable of solubilising P. The P-solubilising bacteria typically outnumber P-solubilising fungi by 2 to 150 fold, though fungal isolates exhibit greater solubilising ability (Gyansehwar et al 2002).
The simplest mechanism of P solubilisation by the microbes is through acidification of the organism’s growth environment. This acidification can simply be a reflection of the nature of the N supply; organisms supplied with N in the growth medium primarily in the ammonium (NH4+) form excrete protons in order to maintain electron neutrality (they have the problem of taking up too many cations – Ca2+, Mg2+, K+, NH4+, and too few anions – SO42-, PO43-, NO3-, and must balance this by pumping out H+). In poorly designed laboratory experiments if N is supplied as ammonium, this results in many organisms being able to solubilise calcium phosphate by this mechanism. This is unlikely to happen in soil because nitrate and not ammonium is the normal source of N for the microbes. The other main P solubilisation strategy is the production and release of organic acids, similar to the process described above for plants. 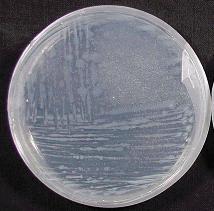 Plate 5. The solubilisation of calcium phosphate (rock phosphate) in agar by micro-organisms is apparent as cleared areas where the organism has been inoculated (as lines) onto this agar plate. This is a simple procedure that has been used to isolate potential solubilisers from soil – a soil sample is diluted in water and streaked onto the calcium phosphate agar media in the plate. The bacteria in the sample which can solubilise the calcium phosphate grow into a colony which then creates a cleared zone in the agar around it as the phosphate source is solubilised.
Plate 5. The solubilisation of calcium phosphate (rock phosphate) in agar by micro-organisms is apparent as cleared areas where the organism has been inoculated (as lines) onto this agar plate. This is a simple procedure that has been used to isolate potential solubilisers from soil – a soil sample is diluted in water and streaked onto the calcium phosphate agar media in the plate. The bacteria in the sample which can solubilise the calcium phosphate grow into a colony which then creates a cleared zone in the agar around it as the phosphate source is solubilised.
Micro-organisms produce a wide range of acids in order to achieve P solubilisation, including acetic, lactic, oxalic, tartaric, succinic, citric, gluconic, ketogluconic, glycolic …… (Gyaneshwar et al 2002). These acids differ greatly in their capacity to solubilise P, and while the extent of desorption/release of P is very soil dependent, it generally requires quite high concentrations of organic acid to achieve this (>100 µM for citric, and > 1mM for oxalic, malic and tartaric).
Of course, production and exudation of organic acids by the micro-organism comes at a substantial energy cost (as discussed above for plants). In order to produce organic acids to excrete into the soil environment, the P-solubilising organism would need a very good supply of energy (for example from root exudates). It’s worth considering an energy budget here. Earlier we considered the simplest system of solubilisation where the plant exudes organic acid into the soil. For the situation where P solubilisation is achieved by a micro-organism living in the vicinity of a plant root (rhizosphere) the energy flow is through plant exudates, taken up by micro-organisms, with part of this energy used for organic acid production. Clearly, this will cost the plant more energy per unit of organic acid released.
Before leaving the rhizosphere its worth considering how long the organic acids will continue to work for, and how large a zone of influence there may be around a P solubilising organism. Organic acids represent an energy source for soil organisms (food for bugs). Studies on the breakdown of organic acids such as citrate and malate added to soil at realistic concentrations similar to rhizosphere concentrations of the order of 10 to 100 µM) show that the acids are rapidly degraded in bulk (non-rhizosphere soil) – the half life is about 2 to 3 h (ie microorganisms will degrade half of the organic acid added at these rhizosphere level concentrations to simpler carbon compounds in 2 to 3 h). In the rhizosphere itself, where there is a much higher population of organisms, degradation will be 2 to 3 times faster (Jones 1998). As for the zone of influence – because organic acids are strongly bound by the soil, they do not move far; the predicted zone of influence for a root is 0.2 to 1.0 mm (Jones 1998). The influence of an individual organism/colony would be a small fraction of this. If we also consider that P is relatively immobile in the soil - clearly, the organism would have to be in the rhizosphere to have any impact on plant growth. Inoculation of the seed, and hence the rhizosphere may work, but treating the bulk soil is very unlikely to be effective.
Field effectiveness of P solubilising organisms. The involvement of micro-organisms in solubilisation of P has been known for more than 100 years, and there is a substantial literature dealing with this issue. However, most research has been at the laboratory culture (petri dish), or glasshouse pot trial scale; the number of field trials is quite small. Unfortunately, there is no simple message from the field trials; some trials showed growth enhancement and/or increased P uptake, but there is large variation in the effectiveness of inoculation with P solubilising organisms (Kucey et al 1989, Gyaneshwar et al 2002). Tandon (1987) undertook a review of this research, and while this report is now 20 years old, the conclusions he reached at that time are still applicable today. Tandon reported that inoculation resulted in 10 to 15% yield increases in 10 out of the 37 experiments he considered; in the remaining trials (70% of cases) there was no increase. Furthermore, he (and subsequent reviewers) considered that even in the trials that showed a yield increase, there was reason to question the validity of the findings. Two of his most important concerns were that:
• In many trials, the inoculation with P-solubilising organisms was not compared to addition of soluble P fertiliser, so there is no direct evidence that plants would respond to increased P availability in these soils. (This is still a valid criticism of recent publications – indeed, some papers provide data to show that the plants do not respond to P fertiliser; i.e. that the soil is not P deficient.)
• The mechanism for plant growth promoting activity of P-solubilising organisms, other than P solubilisation, has not been demonstrated, but has often been claimed. For example, claims that the organisms may produce plant hormones which increase growth. Certainly some P-solubilising organisms do produce plant hormones (e.g. Indole-acetic acid), but the impact of this on plant growth has not been established.
We undertook a rapid review of papers published since Tandon’s 1987 review. On the basis of the number of published papers, research on P-solubilising organisms is concentrated in a limited number of counties (Table 2), with India dominating. Of the field studies published, 10 papers show a yield or biomass increase as a result of inoculation, and 7 show no effect. Of the papers showing a beneficial effect of inoculation with P-solubilising organisms, the benefit ranged from a modest increase (e.g. 10%), to more than two fold increase in one instance. We considered that the results of a further 18 papers could not be reliably interpreted. These papers had one (or several) of three types of limitations.
• The effect of P-solubilising organisms could not be separated from the effect of other beneficial organisms. In several studies using legume test species, a mixed inoculum consisting of rhizobium, P-solubilising organisms, and other organisms considered to be beneficial was applied. A beneficial effect of N supply through nodulation could be expected.
• In many studies no effort was made to establish that the soil was P responsive at all, or within the range of P application used as treatments. In India it is common to compare a “recommended” rate of P fertiliser with a fraction of this rate (e.g. 75%) plus P-solubilising organism inoculation. If these treatments achieve the same yield, the researchers interpret this as a demonstration that P-solubilisation has replaced the remaining (25%) fertiliser. This would only be valid if increasing the fertiliser rate did increase yield, and this was not demonstrated. It may be that 75% of the fertiliser was sufficient to achieve maximum yield, and that the inoculation did nothing.
• A limited number of experiments used treatments of fertiliser, and the same rate of fertiliser plus P-solubilising organism inoculation. If these treatments achieved the same yield, then the researchers interpreted this as a demonstration that P-solubilisation did not occur. Once again, this would only be true if the addition of more P increased plant yield, and this was not demonstrated. It is possible that solubilisation did occur, but the plant was already adequately supplied with P and hence did not grow any better or was limited in its growth by the lack of other nutrients.
Table 2. Origin and nature of research on P-solubilising organisms
|
Country where research was undertaken
|
Total number of papers
|
Number of field studies
|
|
India
|
34
|
16
|
|
China
|
11
|
4
|
|
Brazil
|
9
|
1
|
|
Turkey
|
9
|
5
|
|
Canada
|
8
|
2
|
|
Czechoslovakia
|
7
|
2
|
|
others
|
19
|
5
|
Organisms which are capable of P solubilisation in the laboratory, often fail to achieve this in soil. This can in part be attributed to the more strongly buffered nature of soil systems (relative to laboratory microbial growth media). Organisms which produce acid can solubilise P in poorly buffered media, because the pH is easily lowered by production of H+. However, it requires a great deal more acid production to solubilise P in a buffered soil, and few organisms can achieve this. This is especially true for vertosols, which may contain high levels of lime (calcium carbonate), and are thus able to maintain a constant pH even when relatively large amounts of H+ are added.
We have had the opportunity to test P-solubilising bacteria as part of an ACIAR project we have been undertaking in Madhya Pradesh, India, with scientists from the Indian Institute of Soil Science (IISS). The inoculum used was a mixture of P-solubilising bacteria selected to be effective across a wide range of soil types and crops. This inoculum was developed by scientists at IISS, and is available commercially to farmers in India. Four replicated experiments were undertaken (two experiments in two districts) for two years (2005, 2006). Individual plots were 60 m x 4.5 m. The five treatments applied were inorganic fertiliser at the recommended rate (100%), fertiliser at 75% of the recommended rate (75%), fertiliser at 75% of the recommended rate plus P-solubilising bacteria (75%+PSB), an organic treatment of 8 t/ha of farm yard manure (Org), and this organic treatment plus P-solubilising bacteria (Org+PSB). The recommended P fertiliser rate for the area is 26 kg P/ha, and this was the rate used in the 100% treatment. Other nutrients likely to be limiting plant growth were identified in earlier glasshouse nutrient omission experiments where growth is compared with a complete nutrient addition and this minus the element under test). These nutrients (in this case N, S, and Zn) were added as a basal application to the 100%, 75% and 75%+PSB treatments. The test crop was wheat, grown in the winter or rabi season and the soil at each site was a vertosol. The crops typically received four irrigations. Across all four experiments in each of the two years, there was no significant grain or straw yield increase, or additional P uptake, as a result of the P-solubilising bacteria (Figures 5 and 6). 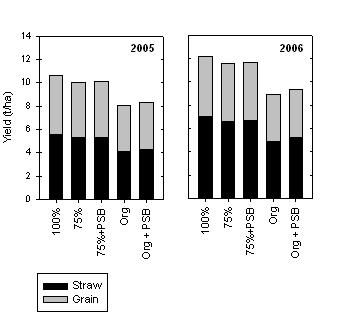 Figure 5. Average grain and straw yields for wheat grown using inorganic fertilization, or organic fertilization, with (+PSB) or without inoculation of seed with P-solubilising bacteria. Yields are the mean of four experiments.
Figure 5. Average grain and straw yields for wheat grown using inorganic fertilization, or organic fertilization, with (+PSB) or without inoculation of seed with P-solubilising bacteria. Yields are the mean of four experiments.
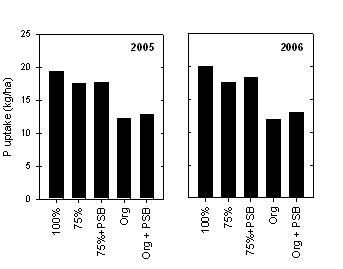 Figure 6. Average P uptake for wheat grown using inorganic fertilization, or organic fertilization, with (+PSB) or without inoculation of seed with P-solubilising bacteria. Yields are the mean of four experiments.
Figure 6. Average P uptake for wheat grown using inorganic fertilization, or organic fertilization, with (+PSB) or without inoculation of seed with P-solubilising bacteria. Yields are the mean of four experiments.
Where does all of this leave us? As with the free-living N2 fixers, we need to consider what benefit could be expected from seed inoculation. As soils contain large numbers of organisms capable of solubilising P, inoculation would only be of benefit if the inoculated strain was much more effective than the organisms already present in the soil. On the basis of the published literature, inoculation does appear to work sometimes, but the circumstances in which this will occur reliably have certainly not been established. At this time we are unable to predict when a positive response will be obtained. A series of trials over different years, in a range of soil types would be needed to establish the reproducibility of the response. Rarely is this done with microbial inoculation experiments. When inoculation with P-solubilising organisms does work, most trials have shown a modest increase in P availability and of crop yield (somewhere in the 10% range). When you consider the substantial energy cost of P solubilisation, a modest increase is probably all you should expect.
How should a farmer respond?
• The “Science” is not complete, so predicting benefits is hard to impossible
• Your situation is unique
• Test the product yourself
• But test it properly – comparison with established alternative; replication.
• Don’t fool yourself.
All easier said than done, but how would you practically go about testing the effectiveness of a microbial inoculant designed to fix N or solubilise P? How would you ensure that you could draw valid conclusions – i.e. not fool yourself? Here are a few simple guidelines that might be useful in formulating your on-farm experiments.
Obviously the work needs to be done in the field, with all the system buffering and spatial variability that brings with it. Replication is therefore essential, as is a random allocation of treatments to strips or plots. Strips are often easier to manage, especially with GPS guidance systems and/or tram tracks upon which to base your treatment application and harvesting. The important thing is to at least match the strip/plot width to that of your harvesting gear, because that is the operation which needs to be easy to do (and do well) to measure the treatment effects. How many times have yields from strips not been collected because harvest was rushed and the contractor didn’t have time to muck around! Having yield monitors on the harvester would also be nice, but you can always fall back on the portable weigh bins if necessary.
The next thing to consider is your reference treatment or control, so that you can interpret the research findings. Ideally it should be something you do currently, and not a ‘nothing applied’ treatment (or not only a nothing applied treatment), as you are generally trying to prove the treatment is as good as, or better than, what you are currently doing (or cheaper). Most people are not (or should not) be growing crops with no inputs at all.
Also, be very clear about what you are trying to test, given the cautionary examples listed earlier. I think most people would accept that many of our current fertiliser use guidelines are best bet options, albeit based on experience gleaned from lots of trials and experience in different farms and soil types. That means that a low pre-plant soil N or P test does not guarantee you a fertiliser response. Make sure that you don’t just have two comparisons in your test – a current practice (eg. your standard rate of starter P) and your biological alternative or treatment of interest. If they produce similar yields you will not be any wiser, as the product could be effective, or the site may not have been responsive in the first place. Having a Nil P treatment in this case will sort that out.
Finally, don’t leap in without giving the product a thorough test in different seasons and paddock conditions. People often question why science takes so long to be sure about something, but the earlier Maxicrop example shows that there will be a range of outcomes with an average effect, and it is important to test often enough to get a realistic estimate of that average effect before you make a change.
References
Dart PJ (1986) Nitrogen fixation associated with non-legumes in agriculture. Plant and Soil 90, 303-334
Edmeades DC (2000) 'Science friction: the Maxicrop case and the aftermath.' (Fertiliser Information Services Ltd, PO Box 9147, Hamilton, New Zealand: Hamilton)
Edmeades DC (2002) The effects of liquid fertilisers derived from natural products on crop, pasture, and animal production: a review. Australian Journal of Agricultural Research 53, 965-976
Einsele, G., C. Limmer, and H.L. Drake. 1996. Non-symbiotic N2-fixation in acidic and pH-neutral forest soils: aerobic and anaerobic differentials. Soil Biology & Biochemistry 28:177-183.
Gyaneshwar, P., G.N. Kumar, L.J. Parekh, and P.S. Poole. 2002. Role of soil microorganisms in improving P nutrition of plants. Plant Soil 245:83-93.
Herridge, D.F., M.B. Peoples, and R.M. Boddey. 2008. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 31:1-18.
Jones, D.L. 1998. Organic acids in the rhizosphere - a critical review. Plant Soil 205:25-44.
Kennedy, I.R., and N. Islam. 2001. The current and potential contribution of asymbiotic nitrogen fixation to nitrogen requirements on farms: a review Australian Journal of Experimental Agriculture 41:447-457.
Lamb, J.A., J.W. Doran, and G.A. Peterson. 1987. Nonsymbiotic dinitrogen fixation in no-till and conventional wheat-fallow systems. Soil Sci Soc Am J 51:356-361.
Lambers, H., F.S. Chapin, and T.L. Pons. 2008. Plant Physiological Ecology. Second Edition. Springer, New York.
Lynch, J.P., and M.D. Ho. 2005. Rhizoeconomics: Carbon costs of phosphorus acquisition. Plant Soil 269:45-56.
Silvester, W., and D. Musgrave. 1991. Free-living diazotrophs, p. 162-186, In M. Dilworth and A. Glen, eds. Biology and Biochemistry of Nitrogen Fixation. Elsevier, Amsterdam.
Smil, V. 1997. Global population and the nitrogen cycle. Scientific American 277:76-81.
Son, Y. 2001. Non-symbiotic nitrogen fixation in forest ecosystems. Ecological Research 16:183-196.
Tandon, H.L.S. 1987. Phosphorus Research and Production in India. Fertiliser Development and Consultation Organization, New Delhi.
Tarafdar, J.C., and A. Jungk. 1987. Phosphatase activity in the rhizosphere and its relation to the depletion of soil organic phosphorus. Biol Fertil Soils 3:199-204.
Triplett, E. 2007. Prospects for significant nitrogen fixation from bacterial endophytes, p. 303-314, In C. Elmerich and W. Newton, eds. Associative and Endophytic Nitrogen-fixing Bacteria and Cyanobacterial Associations. Springer, Dordrecht.
Vitousek, P.M., J.D. Aber, R.H. Howarth, G.E. Likens, P.A. Matson, D.W. Schindler, W.H. Schlesinger, and D.G. Tilman. 1997. Human alteration of the global agricultural cycle: Source and consequences. Ecological Applications 7:737-750.
Contact details
|
Neal Menzies
School of Land, Crop and Food Science, The University of Queensland.
|
Dr Mike Bell
Department of Primary Industries, Queensland
|
Dr Peter Dart
School of Land, Crop and Food Science, The University of Queensland
|
Was this page helpful?
YOUR FEEDBACK